Cette protéine intracellulaire est exprimée dans les tissus hématopoïétiques (le thymus, la rate, la moelle osseuse et les cellules mononucléaires du sang périphérique (PBMCs) comme les cellules T, B, les neutrophiles, les cellules dendritiques, les cellules Killer (NK)). Elle possède, dans sa partie N-terminale, un domaine catalytique (tyrosine phosphatase) et dans sa partie C-terminale, quatre motifs riches en proline, nommés P1, P2, P3 et P4. Le domaine P1 a la capacité de se lier au domaine SH3 (Src homology 3) de la protéine tyrosine kinase Csk. Dans les cellules T, le complexe LYP/Csk est un régulateur négatif de la voie de signalisation TCR. De façon synergique, ce complexe inhibe, selon la déphosphorylation et la phosphorylation de tyrosines ciblées, les kinases de la famille Src comme Lck et Fyn, initiatrices du signal TCR. De plus, LYP peut aussi inhiber l'activation des cellules T, via un nouveau complexe avec le proto-oncogène c-Cbl, en déphosphorylant une autre tyrosine kinase, ZAP-70 [Cloutier and Veillette, 1999,Gjorloff-Wingren et al., 1999](Figure 3.2Représentation schématique des fonctions connues et putatives de PTPN22 dans la voie de signalisation du récepteur de surface des lymphocytes T (TCR)).
Les études d'association de différentes maladies auto-immunes précédemment citées ont mis en évidence un rôle du polymorphisme nucléotidique (SNP) faux-sens du gène PTPN22, rs2476601, situé dans le domaine P1 permettant l'interaction avec SH3 de Csk, dans la prédisposition à l'auto-immunité. Ce variant entraîne une substitution de l'acide aminé arginine en tryptophane au niveau du codon 620 (R620W).
Des expériences in vitro ont montré que la présence de ce tryptophane au codon 620 (l'allèle T) provoque une perte de liaison entre les deux protéines LYP et Csk et, par conséquent, suggère une dérégulation de l'activation des cellules T. Les sujets porteurs de cet allèle seraient ainsi enclins à une auto-immunité [Begovich et al., 2004,Bottini et al., 2004].
Bien qu'aucune étude de liaison dans nos familles n'ait identifié un locus majeur de prédisposition au psoriasis sur le chromosome 1p13, l'observation que le psoriasis est médié en partie par les cellules T activées [Lew et al., 2004,Sugiyama et al., 2005] et l'implication du gène PTPN22 dans la dérégulation des lymphocytes T suggèrent que PTPN22 est un bon gène candidat. De nombreuses études ont été réalisées pour tester la contribution du variant faux-sens R620W de ce gène PTPN22 à la prédisposition au psoriasis et même à l'arthrite psoriasique. Nistor et ses collaborateurs (2005) ont rapporté une absence d'association de ce variant avec le psoriasis dans 517 familles américaines comprenant 1146 sujets atteints. Ce résultat est confirmé par d'autres études sur des populations diverses soit plus petites, américaines (62 individus atteints), anglaises (279 att.), soit plus grandes, allemandes (793 att.), anglaises (647 att.) [Criswell et al., 2005,Hinks et al., 2005,Huffmeier et al., 2006b,Smith et al., 2008]. Dans le cas de l'arthrite psoriasique, l'association du variant faux-sens est faiblement détectée seulement dans certaines conditions (dépendant de la population ou du sexe) [Hinks et al., 2005,Butt et al., 2006,Huffmeier et al., 2006a]. Cependant, une première étude globale de la région de PTPN22 a été réalisée sur 793 patients allemands, dont la plupart ont développé la maladie avant l'âge de 40 ans (Type I). Une association est montrée entre le psoriasis et un haplotype au niveau de la région 1p13 impliquant quatre TagSNPs, trois étant situés dans le gène RSBN1 (round spermatid basic protein 1 gene) et un dans le gène FLJ22588, gènes localisés de part et d'autre de PTPN22 (Pc=0.02) [Huffmeier et al., 2006b]. De plus, une étude cas-témoins sur 900 patients anglais révèle également une association avec des polymorphismes de cette région et le psoriasis de type I. Une association significative est observée entre le psoriasis et deux variants, rs1217414 et rs3789604, localisés dans l'intron 1 du gène PTPN22 et l'exon 1 du gène RSBN1 (P=0.003 et P=0.0002, respectivement) [Smith et al., 2008]. L'ensemble de ces associations sont indépendantes de l'allèle à risque HLA-Cw6 [Huffmeier et al., 2006b,Smith et al., 2008]. L'association du psoriasis avec rs3789604 a été confirmée lors d'une méta-analyse sur 4811 cas et 5662 témoins (P=3.45x10![]() ) [Li et al., 2008]. Ce SNP est particulièrement intéressant car il a tout d'abord été associé avec RA, de manière indépendante de l'association bien caractérisée avec R620W [Carlton et al., 2005]. Ce variant synonyme est localisé à 1496 pb en amont du gène PTPN22 dans l'exon 1 du gène RSBN1. Il semble peu probable que ce gène puisse être un gène de susceptibilité au psoriasis car ce gène semble avoir un rôle important dans la spermatogenèse (différenciation des spermatides en spermatozoïdes), et dans la régulation transcriptionnelle de gènes dans les cellules germinales haploïdes (les spermatides) [Takahashi et al., 2004]. Néanmoins, le polymorphisme pourrait jouer un rôle dans l'activation des cellules T car il est prédit qu'il se situe dans un site de liaison pour différents facteurs de transcription, spécialement pour deux facteurs, Pax-5 et Pax-4, qui pourraient réguler l'expression du gène PTPN22. De plus, PAX5 code pour une protéine activatrice spécifique des cellules B. En absence de PAX5, les cellules pro-B peuvent se différencier, selon les stimuli, en d'autres cellules immunitaires telles que les cellules T, les macrophages ou les cellules dendritiques (cellules présentatrice d'antigènes aux lymphocytes) [Smith et al., 2008].
) [Li et al., 2008]. Ce SNP est particulièrement intéressant car il a tout d'abord été associé avec RA, de manière indépendante de l'association bien caractérisée avec R620W [Carlton et al., 2005]. Ce variant synonyme est localisé à 1496 pb en amont du gène PTPN22 dans l'exon 1 du gène RSBN1. Il semble peu probable que ce gène puisse être un gène de susceptibilité au psoriasis car ce gène semble avoir un rôle important dans la spermatogenèse (différenciation des spermatides en spermatozoïdes), et dans la régulation transcriptionnelle de gènes dans les cellules germinales haploïdes (les spermatides) [Takahashi et al., 2004]. Néanmoins, le polymorphisme pourrait jouer un rôle dans l'activation des cellules T car il est prédit qu'il se situe dans un site de liaison pour différents facteurs de transcription, spécialement pour deux facteurs, Pax-5 et Pax-4, qui pourraient réguler l'expression du gène PTPN22. De plus, PAX5 code pour une protéine activatrice spécifique des cellules B. En absence de PAX5, les cellules pro-B peuvent se différencier, selon les stimuli, en d'autres cellules immunitaires telles que les cellules T, les macrophages ou les cellules dendritiques (cellules présentatrice d'antigènes aux lymphocytes) [Smith et al., 2008].
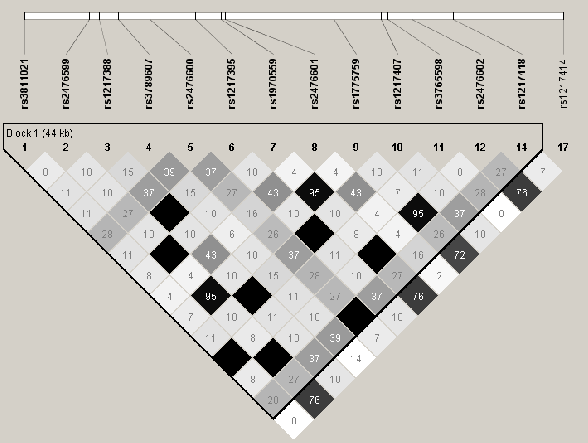
Lorsque nous avons entrepris l'étude de ce gène, en 2005, la majorité de ces études ne prenaient en compte qu'un seul variant, et une association entre le gène PTPN22 et le psoriasis ne pouvait donc pas être exclue formellement. Nous avons donc entrepris une approche systématique 'Tagging SNP' pour tester les gènes candidats. Dans le cas du gène PTPN22, nous avons donc voulu savoir si un variant, autre que R620W, pouvait être associé au psoriasis. Pour ce faire, une étude systématique de PTPN22 a été réalisée et d'après l'étude du déséquilibre de liaison au niveau du gène, 5 TagSNPs, dont rs2476601, ont été testés (Table 3.5Identification et analyse d'association des TagSNPs de PTPN22 sur 45 familles (Lot I) par FBAT, Figure 3.3 Structure DL du gène PTPN22). Les résultats avec le programme FBAT pour les 5 variants de PTPN22 sont présentés dans le tableau 3.5Identification et analyse d'association des TagSNPs de PTPN22 sur 45 familles (Lot I) par FBAT.
Cette analyse a suggéré une association possible entre le psoriasis et deux SNPs introniques, rs2476599 (P=0.08) et rs1217414 (P=0.07), dont le dernier SNP ayant été déjà décrit comme étant associé au psoriasis [Smith et al., 2008]. Cependant, cette tendance n'a pas été confirmée par la suite en analysant les haplotypes constitués par les 5 SNPs dans le Lot I par FBAT (Table 3.6Analyse d'association des haplotypes de PTPN22 sur le Lot I par FBAT) ou en analysant deux SNPs dans le Lot I par la méthode "LNMs" (Table 3.7Analyse d'association des TagSNPs de PTPN22 sur le Lot I avec la méthode "LNMs") ou encore en étudiant ces derniers dans le Lot II ou les deux lots combinés par FBAT (Table 3.8Analyse d'association des TagSNPs de PTPN22, suggestifs dans le Lot I, faite sur un deuxième lot de 83 familles (Lot II) et sur l'ensemble des familles (Lot I et Lot II)).
De plus, aucun des 5 TagSNPs ne montre une association avec le psoriasis après stratification des familles du Lot I selon l'haplotype HLA-Cw6 (Table 3.9Analyse d'association des TagSNPs de PTPN22 dans les sous groupes porteurs ou non de l'allèle à risque HLA-Cw6 dans le Lot I).
Ce manque d'interaction entre les deux loci dans la prédisposition au psoriasis a également été décrit dans d'autres études allemandes et anglaises [Huffmeier et al., 2006b,Smith et al., 2008].
En revanche, l'absence d'association entre le polymorphisme faux-sens R602W de PTPN22 (rs2476601) largement étudié dans différentes maladies auto-immunes et le psoriasis montrée par nos différentes analyses est en accord avec les nombreuses études réalisées sur le psoriasis [Criswell et al., 2005,Hinks et al., 2005,Nistor et al., 2005,Huffmeier et al., 2006b,Lee et al., 2007,Smith et al., 2008].
| Lot I |
||||||
| rs2476599 | G>A | 27.1 | 0.45 | 0.93 [0.76-1.13] | 0.86 [0.70-1.05] | |
| rs1217414 | C>T | 27.8 | 0.18 | 0.86 [0.69-1.07] | 0.74 [0.59-0.92] | |
|
||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Lot I | |||||||||
| Stratifié selon: | |||||||||
| présence HLA-Cw6 | absence HLA-Cw6 | ||||||||
| MAF | P | N | Z | P | N | ||||
| rs3811021 | T>C | 23 | 0.68 | 36 | -0.498 | 0.62 | 37 | 1.222 | 0.22 |
| rs2476599 | G>A | 27.1 | 0.08 | 52 | -0.916 | 0.36 | 52 | -1.538 | 0.12 |
| rs1217388 | A>G | 21.5 | 0.57 | 49 | 0.745 | 0.46 | 33 | -0.052 | 0.96 |
| rs2476601 | G>A | 7.3 | 0.64 | 23 | -0.845 | 0.40 | 19 | 0.252 | 0.80 |
| rs1217414 | C>T | 27.8 | 0.07 | 52 | -1.127 | 0.26 | 51 | -1.346 | 0.18 |
L'ensemble de ces résultats ne montre pas de rôle significatif du gène PTPN22 dans la prédisposition au psoriasis dans nos familles. Une association, peu significative, a été cependant observée avec un variant présent dans l'intron 1 du gène PTPN22 (rs1217414) dans notre premier lot de familles. Par ailleurs, la région PTPN22, ainsi que ce variant, qui peut avoir un effet sur la transcription du gène, ont été montrés récemment associés au psoriasis, particulièrement de type I [Huffmeier et al., 2006b,Smith et al., 2008]. Dans notre étude, l'âge moyen du début des symptômes chez les patients des familles du premier lot est de 19 ans (psoriasis de type I). Ceci pourrait expliquer la tendance d'association observée pour ce marqueur. Le second variant (rs3789604) récemment décrit comme associé au psoriasis, qui semble pouvoir modifier la fixation de certains facteurs de transcription, n'a pas été étudié lors de notre étude. Contrairement aux deux études récentes, nous nous sommes, en effet, focalisés sur le gène lui-même et non sur la région environnante [Smith et al., 2008].
Malgré le manque d'association avec l'un des variants étudiés (rs3811021) qui est en fort déséquilibre de liaison (r![]() =0.89) avec le second variant associé (données non montrées), il serait néanmoins intéressant d'étudier ce variant dans nos familles du lot I car il pourrait être associé à la maladie, indépendamment du variant rs3811021, comme observé dans l'étude initiale [Smith et al., 2008].
Notre étude confirme que le polymorphisme R620W n'est pas associé au psoriasis, répliquant ainsi les résultats de nombreuses études.
=0.89) avec le second variant associé (données non montrées), il serait néanmoins intéressant d'étudier ce variant dans nos familles du lot I car il pourrait être associé à la maladie, indépendamment du variant rs3811021, comme observé dans l'étude initiale [Smith et al., 2008].
Notre étude confirme que le polymorphisme R620W n'est pas associé au psoriasis, répliquant ainsi les résultats de nombreuses études.
En conclusion, même s'il existe peu de preuves d'association du gène PTPN22 avec le psoriasis dans nos familles, des études récentes montrent une association de la région avec la maladie. Or, cette région est présente dans une région de fort déséquilibre de liaison contenant 6 gènes, mais seul PTPN22 pourrait avoir de par sa fonction un rôle dans le psoriasis. Ainsi, afin d'exclure ou non ce gène comme gène prédisposant au psoriasis dans la population française, d'autres études (par exemple, au niveau de la région promotrice (rs3811021)) sont nécessaires.