Le gŤne FLG
La filagrine, codťe par le gŤne FLG, est une protťine essentielle de la barriŤre ťpidermique au niveau de la couche terminale de la peau, la couche cornťe. Elle provient du clivage en 10-12 protťines d'une grande protťine (>400kDa), la profilagrine, constituant important des granules de kťratohyaline prťsents dans la couche granuleuse de l'ťpiderme [Candi et al., 2005]. Lors du stade terminal de la diffťrenciation des kťratinocytes, la profilagrine est transformťe en filagrine, au niveau de la couche cornťe, formant la matrice cytoplasmique des cornťocytes, qui sont les cellules anuclťťes propres de cette couche. La filagrine est capable d'agrťger les filaments du cytosquelette des kťratinocytes pour mettre en place une matrice dense, rťticulťe par l'action des transglutaminases, et former l'enveloppe cellulaire cornifiťe, essentielle dans la protection de la peau.
Des allŤles de type de "perte de fonction" de ce gŤne, R501X et 2282del4, frťquents dans la population europťenne ( 4% et 1% respectivement), ont ťtť rťcemment identifiťs comme ťtant la cause d'une maladie de peau monogťnique, l'ichtyose vulgaire [Sandilands et al., 2006,Smith et al., 2006]. Ce gŤne reprťsente ťgalement un facteur de prťdisposition ŗ une maladie de peau ŗ hťrťditť complexe, la dermatite atopique (AD), et plus particuliŤrement l'AD associť ŗ des problŤmes allergiques et ŗ de l'asthme [Palmer et al., 2006,Palmer et al., 2007]. Cette derniŤre association a ťtť rťpliquťe dans de nombreuses ťtudes faites sur des populations europťennes et japonaises montrant ainsi la robustesse de cette association [Marenholz et al., 2006,Ruether et al., 2006,Weidinger et al., 2006,Barker et al., 2007,Stemmler et al., 2007,Nomura et al., 2008]. Ces variants provoquent une traduction prťmaturťe de la profilagrine au niveau de la sťquence codant pour la premiŤre filagrine et entraÓnent donc une perte de la production de la filagrine et une formation altťrťe de la barriŤre ťpidermique [Smith et al., 2006]. D'autres variants codants semblent intervenir dans la pathogenŤse de l'AD tels que le variant rare 3702delG qui provoque la fin de la traduction du profilagrine au niveau du domaine formant la troisiŤme filagrine [Sandilands et al., 2006].
Il existe un chevauchement de plusieurs loci de prťdisposition potentiels entre deux maladies complexes de peau, le psoriasis et l'AD, sur les chromosomes 3q21, 17q25, 20p12 et surtout, sur le chromosome 1q21 (PSORS4 pour le psoriasis) au niveau du complexe de diffťrenciation ťpidermique (EDC) qui regroupe plusieurs gŤnes codant des protťines de structure trouvťes dans la couche externe de l'ťpiderme diffťrenciťe dont la filagrine [Bowcock and Cookson, 2004]. De plus, une des caractťristiques du psoriasis est une altťration de la diffťrenciation des kťratinocytes et une inflammation cutanťe. Histologiquement, il est observť que la couche granuleuse est rťduite ou absente, et/ou des changements dans l'expression de plusieurs protťines du complexe EDC dans l'ťpiderme lťsť chez les patients atteints de psoriasis, comparťe ŗ l'ťpiderme non lťsť ou ŗ celui d'un individu sain. Plus particuliŤrement, la filagrine n'est peu ou pas exprimťe dans la couche granuleuse dans l'ťpiderme lťsť, contrairement ŗ celle de l'ťpiderme non lťsť [Huffmeier et al., 2007]. Il est donc intťressant de voir si des altťrations dans ce gŤne, pouvant jouer sur l'expression de celui-ci, pourraient contribuer ŗ la pathogenŤse du psoriasis. Trois ťtudes ont d'ailleurs ťtť rťalisťes afin de tester la contribution de deux variants avec perte de fonction ("variants nuls") du gŤne FLG, R501X et 2282del4, ŗ la prťdisposition au psoriasis. HŁffmeier et ses collaborateurs (2007) ont rapportť un manque d'association de ces variants avec la maladie lors de l'ťtude cas-tťmoins de 375 sujets allemands atteints de psoriasis vulgaris et aussi de 375 sujets allemands atteints de psoriasis arthritique. Ces rťsultats ont ťtť confirmťs lors de l'analyse de 271 patients irlandais et de 420 patients anglais atteints de psoriasis (analysťs sťparťment ou de faÁon combinťe) ainsi que lors de l'analyse de 360 patients allemands (versus 2117 et 276 tťmoins) [Zhao et al., 2007,Weichenthal et al., 2007].
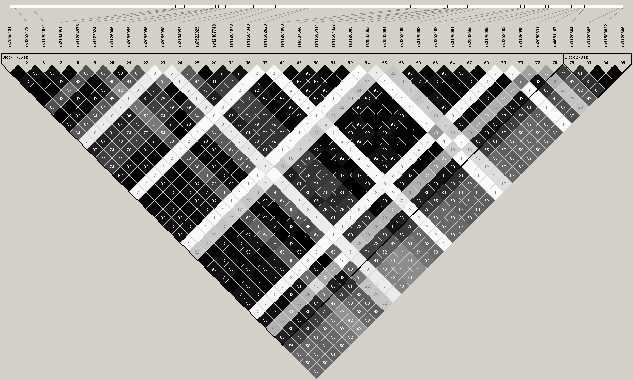
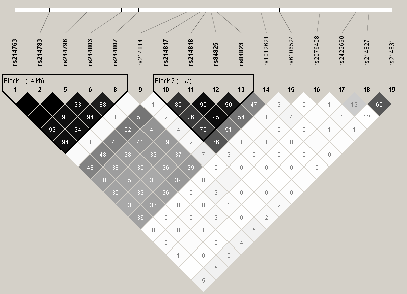
Cependant, ces ťtudes ne prennent en compte que les deux variants nuls du gŤne associť ŗ l'AD et ŗ l'ichtyose vulgaire. Nous avons donc entrepris une approche systťmatique 'Tagging SNP' pour tester ce gŤne. Nous avons voulu tester si un autre variant, prťsent dans la partie codante, non codante ou dans la partie rťgulatrice (+20kb), pouvait Ítre responsable dans la pathogenŤse du psoriasis. En plus des trois variants prťdisposant aux maladies citťes plus haut (3720delG, 2282del4 et R501X), nous avons sťlectionnť 7 SNPs supplťmentaires, d'aprŤs la structure DL du gŤne FLG (Figure 3.14Structure DL du gŤne FLG, Table 3.42Identification et analyse d'association des TagSNPs de FLG sur le Lot I par FBAT). Pour des raisons techniques, deux variants 2282del4 et 3720delG n'ont pas pu Ítre gťnotypťs. L'analyse par FBAT du lot I suggŤre une association, bien que non significative, entre le variant R501X (exon 3, P=0.07) pour lequel des associations ont dťjŗ ťtť rapportťes dans la littťrature et le psoriasis. Notre ťtude rťvŤle une association statistiquement significative entre la maladie et rs1858483, un variant prťsent en amont du gŤneFLG (P=0.001). Ce rťsultat n'est pas confirmť avec la mťthode "LNMs" (Table 3.43Analyse d'association des TagSNPs de FLG dans le Lot I avec la mťthode "LNMs" ). L'analyse haplotypique du gŤne FLG, dans le lot I, a ťtť rťalisťe par l'ťtude des combinaisons de paires de SNPs et indique une association avec la maladie d'une majoritť d'haplotype ŗ effet protecteur (Z<0) avec un P, aprŤs 1 000 000 permutations, variant de 0.0009 et 0.04 (Table 3.44Analyse d'association des haplotypes de FLG sur le Lot I par FBAT). De plus, les haplotypes
| Lot I | ||||||
| MAF | P | Z | ||||
| 1/11 | GT | 10.4 | -3.068 | 0.0022 | 0.0023 | (0.019) |
| 1/12 | GG | 10.5 | -2.092 | 0.036 | 0.037 | (0.11) |
| 2/11 | CT | 10.2 | -2.614 | 0.0090 | 0.011 | (0.022) |
| GC | 75.1 | 2.033 | 0.042 | 0.039 | 2(0.0042) | |
| GT | 10.1 | -3.120 | 0.0018 | 0.0016 | ||
| 4/12 | GG | 11.3 | -2.017 | 0.044 | 0.052 | (0.14) |
| CC | 87.8 | 3.978 | 0.000069 | 0.000066 | 2(0.000063) | |
| CT | 10.8 | -3.355 | 0.00079 | 0.00086 | ||
| AC | 79.7 | 2.467 | 0.014 | 0.013 | 2(0.013) | |
| AT | 10.4 | -2.721 | 0.0065 | 0.0041 | ||
| 10/11 | GT | 10.1 | -2.806 | 0.0050 | 0.0055 | (0.053) |
| 10/12 | GG | 10.0 | -2.089 | 0.037 | 0.039 | (0.25) |
| CT | 71.4 | 2.277 | 0.023 | 0.024 | 2(0.0098) | |
| TT | 10.2 | -3.018 | 0.0025 | 0.0026 | ||
| CA | 73.7 | 2.276 | 0.023 | 0.027 | 2(0.019) | |
| TA | 10.5 | -2.689 | 0.0072 | 0.0095 | ||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Lot I | |||||||||
| Stratifiť selon: | |||||||||
| prťsence HLA-Cw6 | absence HLA-Cw6 | ||||||||
| MAF | P | N | Z | P | N | ||||
| rs12730241 | G>A | 14.2 | 0.31 | 45 | 0.895 | 0.37 | 32 | 0.429 | 0.67 |
| rs2065955 | C>G | 12.8 | 0.68 | 43 | 0.319 | 0.75 | 28 | 0.236 | 0.81 |
| rs7522925 | G>A | 11.5 | 0.53 | 42 | 0.739 | 0.46 | 25 | 0.031 | 0.98 |
| FLG_R501X | C>T | 1.7 | 0.07 | 9 | *** | *** | 5 | *** | *** |
| rs11582620 | A>G | 9.7 | 0.62 | 29 | 0.000 | 1.0 | 25 | -0.703 | 0.48 |
| rs1933064 | G>A | 42.9 | 0.17 | 58 | -0.792 | 0.43 | 52 | 2.793 | 0.005 |
| rs1858483 | C>T | 13.1 | 0.001 | 26 | -2.300 | 0.02 | 35 | -2.226 | 0.02 |
| rs2050631 | A>G | 25.6 | 0.22 | 52 | -0.739 | 0.46 | 45 | -0.910 | 0.36 |
| rs3120664 | T>G | 15.6 | 0.93 | 42 | 0.269 | 0.79 | 28 | -0.196 | 0.84 |
| rs3120665 | A>G | 13.2 | 0.93 | 39 | -0.030 | 0.98 | 27 | -0.092 | 0.93 |
associťs sont une combinaison avec au moins un de ces SNPs R501X, rs1858483 et rs2050631, pour lesquels une association est suggťrťe (sauf pour rs2050631) avec la maladie dans l'ťtude individuelle des SNPs. Le meilleur rťsultat est obtenu par la combinaison des deux SNPs associťs individuellement, R501X et rs1858483 : l'haplotype ŗ risque, CC (88%, Z=3.98, P=0.00007) et inversement, l'haplotype protecteur, CT (11%, Z=-3.36, P=0.0009). Ce dernier semble reflťter le rťsultat obtenu lors du test du SNP rs1858483 dans le lot I. De plus, l'ťtude du lot II seul n'est certes pas significative mais l'analyse des deux lots montre encore une association du marqueur rs18585483 avec le psoriasis (P=0.009) (Table 3.45Analyse d'association des TagSNPs de FLG, suggestifs dans le Lot I, faite sur le Lot II et ainsi que sur l'ensemble des lots (Lot I et Lot II)). Pour le marqueur rare R501X, le lot II s'avŤre peu informatif pour permettre une confirmation. Lors de la derniŤre ťtude, l'association de rs1858483 dans le lot I semble Ítre indťpendante du paramŤtre "porteur" ou "non porteur" de l'allŤle HLA-Cw6 avec un P=0.02 pour les deux sous-groupes (Table 3.46Analyse d'association des TagSNPs deFLG dans les sous groupes porteurs ou non de l'allŤle ŗ risque HLA-Cw6 dans le Lot I). Un autre marqueur, rs1933064, paraÓt Ítre associť au psoriasis lorsque l'on ne considŤre que les individus ne portant pas l'allŤle ŗ risque.
Ces rťsultats semblent indiquer un rŰle dans la maladie d'un SNP, localisť en amont du gŤne filagrine dans le psoriasis, agissant indťpendamment du locus PSORS1, mais ne confirme pas celui de R501X. Dans notre population franÁaise, la non-contribution de ce variant nul dans la prťdisposition au psoriasis s'avŤre Ítre en accord avec les ťtudes indťpendantes faites par quatre ťquipes europťennes et chinoises [Huffmeier et al., 2007,Zhao et al., 2007,Chang et al., 2008b,Weichenthal et al., 2007]. De plus, dans notre ťtude sur la filagrine, nous nous sommes intťressťs ŗ la globalitť du gŤne en tenant compte de la structure d'au moins 20kb en amont et en aval du gŤne. Il existe 10 variants codants et validťs dans la population caucasienne. Quatre de ces dix SNPs ont ťtť testťs dans cette ťtude et les 6 autres sont en fort dťsťquilibre de liaison avec les 4 analysťs. En raison de l'absence d'association avec les SNPs codants connus pouvant intervenir dans la conformation de la protťine et l'association d'un marqueur en amont du gŤne avec le psoriasis, la rťgion devra Ítre approfondie pour ťlucider la nature exacte du variant causal. Ceci est d'autant plus intťressant qu'un des variants codants (P478S) est associť au psoriasis dans la population chinoise (P=0.007) [Chang et al., 2008b]. Il est ťgalement possible que ce soit un SNP inconnu dans les bases de donnťes en dťsťquilibre de liaison avec ceux associťs, codants ou non. Par exemple, les trois SNPs de la filagrine associťs aux maladies de peau, R501X, 2282del4 et3720delG n'ťtaient pas mentionnťs dans la base de donnťe NCBI. On peut aussi supposer que le variant associť est en dťsťquilibre de liaison avec un autre provenant d'un gŤne proche. Ceci serait intťressant dans le cas de la filagrine car il existe, en effet, un autre gŤne de la mÍme famille, trŤs proche du gŤne FLG nommťe FLG2, et le marqueur associť se situe entre les deux gŤnes. Cependant, d'aprŤs la structure en blocs DL avec les donnťes HapMap (non montrťes), les
deux gŤnes ne paraissent pas en fort dťsťquilibre de liaison.
Les gŤnes TGM1, TGM3, TGM5 et TGM6
Comme discutť prťcťdemment, la fonction de barriŤre de l'ťpiderme est assurťe en surface par une structure produite par les cornťocytes appelťe la couche cornťe. Cette couche cornťe est composťe de nombreuses protťines (comme la filagrine), rťticulťes par une famille d'enzymes, les transglutaminases. Ces derniŤres catalysent des rťactions de pontage protťiques, formant ainsi un complexe protťique macromolťculaire particuliŤrement stable et insoluble. Ces enzymes ont donc un rŰle important dans la diffťrenciation terminale de l'ťpiderme.
Des mutations de type "perte de fonction" dans deux gŤnes codant pour les transglutaminases 1 et 5 (TGM1 et TGM5) ont ťtť identifiťes comme ťtant la cause de trois ťrythrodermies ichtyosiformes congťnitales rares rťcessives autosomales de la peau, l'ichtyose lamellaire, l'ichtyose non bulleuse et la forme acral du syndrome de desquamation continue [Russell et al., 1995,Cassidy et al., 2005]. Ce sont des dťsordres de cornification, caractťrisťs par une sťcheresse cutanťe prononcťe au niveau du tronc, des rťgions proximales des membres supťrieurs, du cuir chevelu et du cou et un ťpaississement majeur de l'ťpiderme. Le syndrome de desquamation continue acral ou 'peeling skin syndrome' est caractťrisť par une desquamation continue de la couche cornťe prťdominante au niveau du dos des mains et des pieds.
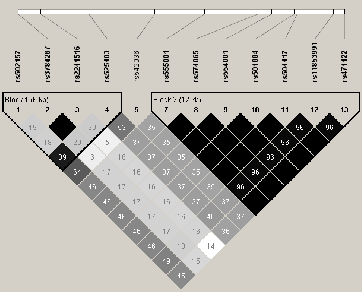
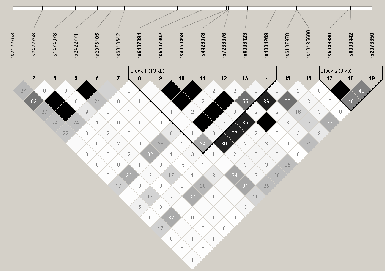
Trois gŤnes codants pour les transglutaminases 1, 3 et 6 (TGM1, TGM3 et TGM6) sont situťs sur les chromosomes 14q et 20p, deux des loci de prťdisposition dans nos 45 familles ťtudiťes. Nous nous sommes donc intťressťs ŗ ces 4 gŤnes pouvant, de par leur fonction ou leur position sur le gťnome, jouer un rŰle dans une ťrythrodermie ťrythťmato-squameuse telle que le psoriasis. L'ťtude du dťsťquilibre de liaison dans les 4 gŤnes a permis de sťlectionner 5 TagSNPs pour TGM1, 9 pour TGM3, 7 pour TGM5 et 11 pour TGM6 (Figures 3.15Structure DL du gŤne TGM3,3.16Structure DL du gŤne TGM5 et 3.17Structure DL du gŤne TGM6). La structure DL du gŤne TGM1 n'est pas prťsentťe car il n'existait ŗ l'ťpoque dans la base HapMap qu'un seul SNP de frťquence supťrieure ŗ 5%. L'ensemble des informations sur les marqueurs choisis est exposť dans le tableau 3.47Identification et analyse d'association des TagSNPs de TGM1, TGM3, TGM5 et TGM6 sur le Lot I par FBAT. L'analyse des TagSNPs par FBAT dans les 45 grandes familles ne montre aucun rťsultat significatif. Cependant, lors de l'analyse des haplotypes pour chaque gŤne, une association faible entre le psoriasis et un haplotype rare de 6.5% de TGM5 (ACCATGG) est observťe avec une valeur de P de 0.04 (basťe sur 1000000 permutations) (Tableau 3.48Analyse d'association des haplotypes de TGM1, TGM3, TGM5 et TGM6 sur le Lot I par FBAT). AprŤs la stratification des familles selon l'haplotype ŗ risque de PSORS1, l'analyse des TagSNPs rťvŤle des associations entre trois variants introniques (rs2241516, rs11070398 et rs542036) et la maladie (P=0.001, P=0.002, P=0.02 respectivement) (Tableau 3.49Analyse d'association des TagSNPs de TGM1, TGM3, TGM5 et TGM6 dans les sous groupes porteurs ou non de l'allŤle ŗ risque HLA-Cw6 dans le Lot I). L'association avec ces SNPs
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Lot I | ||||
| MAF | ||||
| H1 | TGCCA | 60 | 0.83 | |
| H2 | TGCAA | 12.6 | ||
| H3 | CCTCA | 10 | ||
| H4 | TGCAC | 8.6 | ||
| H5 | TGCCC | 6.3 | ||
| H1 | TGAACA | 50.3 | 0.78 | |
| H2 | TGAACC | 10.9 | ||
| H3 | TGAATA | 7.2 | ||
| H4 | CGGGCA | 6.7 | ||
| H5 | CGGGTA | 5 | ||
| H1 | TGAACTAG | 41.3 | 0.32 | |
| H2 | TGAACTGA | 11.6 | ||
| H3 | TGAACCGA | 10.9 | ||
B.Association
| Lot I | |||||||
| MAF | Z | P | |||||
| H1 | CGTGTTG | 54.9 | -0.054 | 0.96 | 0.98 | 4(0.77) | |
| H2 | AGTACGG | 14.4 | -0.332 | 0.74 | 0.74 | ||
| H3 | CGTGTGG | 7.5 | -0.774 | 0.44 | 0.40 | ||
| H4 | ACCATGG | 6.5 | 2.121 | 0.03 | 0.04 | ||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |